Lexikon der Biologie: Algen
Algen [von latein. algae = diverse Sippen von Seegras und Tang], Phycophyta, nach der bisherigen Systematik Abteilung des Pflanzenreichs mit (je nach zugrundegelegter Systematik) 12–15 Klassen ( vgl. Tab. ); nach der neueren Systematik ein Organisationstyp des Pflanzenreichs mit 9 Abteilungen, die vielfach den früheren Klassen entsprechen. Algen sind kernhaltige, eukaryotische, photoautotrophe (selten mixotrophe) ein- oder vielzellige, verschieden gefärbte Thallophyten. Die aus traditionellen Gründen oft noch als Blaualgen bezeichneten Cyanobakterien besitzen wie die Algen eine oxygene Photosynthese, sind aber prokaryotisch (Prokaryoten) organisiert. Die Sporenbehälter der Algen (Sporangien) sind stets, ihre Geschlechtsorgane in der Regel einzellig (Gametangien: Spermatogonien mit Spermatozoiden bzw. Spermatangien mit Spermatien sowie Oogonium bzw. Karpogon). Die Plastiden nahezu aller Algen enthalten Chlorophyll a (Chlorophylle), daneben meist noch eine weitere Chlorophyllkomponente und akzessorische Pigmente, wie Phycobiline (Phycobiliproteine), Carotine und Xanthophylle (Carotinoide); es gibt jedoch auch einige wenige, sekundär chlorophyllos gewordene Arten. Die meisten Algen sind an das Leben im Wasser gebunden.
Über ihre stammesgeschichtliche Entwicklung weiß man mangels geringer fossiler Funde wenig (Archäophytikum). Bislang nahm man an, daß es erst vor 1 Milliarde Jahren mehrzellige eukaryotische Algen gab. 1995 fanden chinesische Wissenschaftler in 1,7 Milliarden Jahre alten Gesteinsschichten Abdrücke von bis zu 3 cm großen Algen, die Verwandte der heutigen Braunalgen zu sein scheinen. Die eukaryotischen Algen (Algen im eigentlichen Sinne) sind entstanden, indem eine heterotrophe eukaryotische Zelle einen Cyanobakterien-ähnlichen Prokaryoten aufgenommen (primäre Endocytobiose; Endosymbiontenhypothese) und ihn auf die Stufe eines Organells reduziert hat. Hierfür sprechen eine Reihe morphologischer Merkmale, sowie biochemische und molekulare Eigenschaften der Plastiden. Anhand der Plastiden-Morphologie kann man die Algen in solche einteilen, deren Plastiden von 2 Membranen umhüllt sind, und in solche mit 3 bzw. 4 Hüllmembranen. 2 umhüllende Membranen finden sich bei den Glaucophyta, Chlorophyta und Rhodophyta (wie auch bei den Chloroplasten der Landpflanzen), während 3 Membranen bei den Dinophyta und Euglenophyta (Euglenozoa), 4 bei den Haptophyta, Heterokontophyta, Cryptophyta und Chlorarachniophyta vorkommen. Heute weiß man, daß Algen mit zwei Plastidenmembranen in primärer Endocytobiose entstanden sind, während Algen mit Plastiden mit mehr als 2 Hüllmembranen durch die Aufnahme einer bereits plastidenhaltigen eukaryotischen Zelle in eine andere eukaryotische Zelle entstanden sind (sekundäre Endocytobiose). Dies wird unter anderem durch das Vorkommen eines zwischen der zweiten und dritten Membran gelegenen Restzellkerns (Nucleomorph) bei den Cryptophyta und Chlorarachniophyta belegt. Es wird angenommen, daß der Vorgang der primären Endocytobiose monophyletisch ist, während sekundäre Endocytobiosen polyphyletisch mindestens sechsmal unabhängig etabliert wurden. Die einzelnen Algen-Abteilungen haben sich vermutlich schon früh separiert und seit langem eine getrennte Entwicklung durchlaufen. Heute kennt man ca. 39 000 beschriebene Arten, der gesamte Artenbestand wird auf bis zu 280 000 Arten geschätzt. Die Formenvielfalt reicht von 1 μm großen Einzellern bis hin zu meterlangen Großtangen.
Die Fortpflanzungszellen der Algen (Gameten, Sporen) sind meist begeißelt; der Geißelbau ist gleich dem anderer Eukaryoten. Oft werden 2 Geißeln ausgebildet, die nach vorn (Zuggeißel), seltener nach hinten (Schubgeißel und Schleppgeißel) gerichtet sind. Sie können gleich (isokont) oder ungleich lang (heterokont) sein, dabei glatt und peitschenartig auslaufen (Peitschengeißel) oder mit Flimmerhaaren (Flimmergeißel) besetzt sein. Viele einzellige Algen haben keine feste Zellwand, sondern besitzen nur eine feste, periphere, proteinreiche cytoplasmatische Außenschicht (Pellicula, Periplast). Die meisten Algenzellen sind von einer festen Zellwand umgeben, bestehend aus einer gelartigen, leicht verschleimenden, nicht kristallinen Grundsubstanz (meist Pektin), die in ein kristallines, meist fibrilläres Grundgerüst eingelagert ist. Strukturbildende Makromoleküle des Grundgerüstes sind überwiegend Cellulose, seltener Xylane oder Mannane. Häufig sind die Zellwände durch Inkrustierung mit amorpher Kieselsäure (Kieselalgen) oder Calcium- bzw. Magnesiumcarbonaten (Dasycladales) verhärtet. Viele Algengruppen, wie die Kalkflagellaten und die Corallinaceae, sind daher auch erdgeschichtlich bedeutsame Gesteinsbildner (Kalk). Als weitere Wandsubstanzen können Hemicellulose, Alginsäure (bei Braunalgen) oder Agar (bei Rotalgen) eingelagert sein. Als Reservestoffe werden, häufig im Bereich der Pyrenoide (Chloroplasten), Chrysose, Stärke, Florideenstärke und Paramylon gebildet. – Für die systematische Gliederung der Algen werden neben der Farbstoffzusammensetzung der Plastiden und den gebildeten Reservestoffen ( vgl. Tab. ) auch morphologische, cytologische und entwicklungsgeschichtliche Merkmale benutzt sowie die chemische Zusammensetzung der Zellwände. Das derzeitige System der Algen ist innerhalb der Abteilungen immer noch ein provisorisches System, das in den nächsten Jahrzehnten, unter Berücksichtigung stammesgeschichtlicher Zusammenhänge aufgrund DNA-analytischer Befunde, noch vielfache Änderungen erfahren wird; unter anderem spielen die Plastidenfarbstoffe eine wesentliche Rolle bei der Klassifizierung ( vgl. Tab. ). Innerhalb der Abteilungen erfolgt die weitere Gliederung vielfach nach morphologischen Merkmalen. Dieses Einteilungsprinzip ist im Kern vor allem auf A. Pascher (ehemals Professor in Prag) zurückzuführen. Er stellte unter anderem fest, daß innerhalb einzelner Klassen gleiche morphologische Formen auftreten; darauf begründete er seine Theorie der Parallelentwicklung einzelner, monophyletischer Algenklassen. Danach leiten diese sich von flagellatenähnlichen, monadalen Ausgangsformen ab und erfuhren über mehrere Organisationsstufen eine morphologisch-anatomische Höherentwicklung. Diesen Organisationsstufen kommt vielfach der taxonomische Wert eines Ordnungsmerkmals zu:
1. Rhizopodiale oder amöboide Organisationsstufe: einzellige Algen ohne feste Zellwand und Geißel; bewegen sich amöboid, bilden unter anderem Pseudopodien zur Nahrungsaufnahme; dünne, fadenförmige Fortsätze werden als Rhizopodien bezeichnet.
2. Monadale oder monadoide Organisationsstufe: die Zellen dieser Algen besitzen auch während der vegetativen Phase eine oder mehrere Geißeln und können nach der Zellteilung zu mehrzelligen Kolonien vereinigt bleiben (Coenobium); die Zellen sind von einer Pellicula oder festen Zellwand umhüllt.
3. Kapsale (capsale), tetrasporale oder palmelloide Organisationsstufe: einzellige Algen ohne feste Zellwand und Geißel; stets von einem Gallertmantel eingehüllt; Tendenz zur Koloniebildung.
4. Kokkale (coccale) Organisationsstufe: unbewegliche, unbegeißelte einzellige Algen mit fester Zellwand; Tendenz zur Koloniebildung.
5. Trichale Organisationsstufe: einkernige Zellen zu einfachen oder verzweigten, interkalar oder mit Scheitelzelle wachsenden Fäden vereinigt.
6. Siphonale oder coenoblastische Organisationsstufe: vielkernige (polyenergide), blasen- oder schlauchförmige Algen. Mitunter wird noch eine weitere Organisationsstufe aufgeführt, die aber selten vorkommt, die
7. Siphonocladale Organisationsstufe: sie leitet sich von der siphonalen Organisationsstufe ab, und zwar durch zelluläre Gliederung eines polyenergiden, schlauchförmigen Thallus. Die Zellen dieser Organisationsstufe sind ebenfalls vielkernig.
8. Filz- und Flechtthallus: die Zellfäden bzw. Seitenäste sind verfilzt oder miteinander verflochten, die Zellen untereinander verklebt oder auch verwachsen (Plektenchym).
9. Gewebethallus: höchster Differenzierungsgrad, bei dem die sich multiserial teilenden Zellen in einem Gewebeverband verbunden bleiben. Es bilden sich die den Kormophyten-Organen Wurzel, Sproß und Blatt analogen Rhizoide, Cauloide und Phylloide. Entsprechend ergibt sich durch die verhältnismäßig reiche Zelldifferenzierung eine Untergliederung in Abschluß-, Rinden- und Markgewebe.
Fortpflanzung: Die meisten Algen können sich sowohl vegetativ (asexuell) wie sexuell fortpflanzen. Grundlage der vegetativen Fortpflanzung ist die Mitose. Das hat zur Folge, daß stets erbgleiche Nachkommen entstehen. Die einfachste Form ist die Zweiteilung (Schizotomie). Nach der Mitose erfolgt eine weitgehend äquale Teilung des Protoplasten, so daß aus einer Mutterzelle zwei nahezu gleich große Tochterzellen entstehen. Vielfach bleibt dabei von der Mutterzelle kein Rest übrig, man spricht dann auch von der "potentiellen Unsterblichkeit" dieser Einzeller. Einen größeren Propagationswert hat die Schizogonie. Hierbei laufen in einer Zelle (Sporangium) mehrere mitotische Teilungen ab, und es werden mehrere Tochterzellen (Sporen) gebildet, die nach Aufreißen der Sporangienwand frei werden und heranwachsen. Eine weitere Art der vegetativen Fortpflanzung ist der Zerfall mehrzelliger Algen in wenigerzellige Teile, die ihrerseits durch Zellteilung wieder heranwachsen können. Einige Arten bilden auch Planosporen aus. – Die sexuelle Fortpflanzung teilt sich in die beiden Teilschritte der Gametenverschmelzung (Syngamie) und der Meiose. Je nach Ausbildung der miteinander fusionierenden Gameten (vgl. Abb. bei Befruchtung) unterscheidet man zwischen: a) Isogamie, hierbei sind die Gameten morphologisch gleich gestaltet (können aber trotzdem getrenntgeschlechtlich sein); b) Anisogamie, die beiden begeißelten Gameten sind unterschiedlich groß, und c) Oogamie, die weibliche Geschlechtszelle ist unbeweglich (Eizelle). Bei Einzellern (z. B. Chlamydomonas; Chlamydomonadaceae) können die einzelligen Individuen zu Gameten umgestimmt werden und miteinander verschmelzen (Hologamie). Viele Einzeller und alle Mehrzeller bilden in einer Zelle (Gametangium) mehrere Gameten aus, so daß pro Individuum mehrere und kleinere Geschlechtszellen gebildet werden (Merogamie). Die Syngamie kann folglich als isogame, anisogame oder oogame Merogamie bzw. Hologamie erfolgen. Für das Auffinden der konträrgeschlechtlichen Gameten haben bei vielen Algen die von den weiblichen Individuen abgegebenen Gametenlockstoffe (Sexuallockstoffe mit Pheromon-Charakter; vgl. Tab. ) große Bedeutung. Das Verschmelzungsprodukt zweier Geschlechtszellen ist die Zygote. Sie hat bei Süßwasseralgen häufig die Funktion eines Überdauerungskörpers. Entsprechend dick und widerstandsfähig wird die Zygotenwand ausgebildet. Die Algen können gemischtgeschlechtlich (monözisch) sein, d. h., auf dem gleichen Thallus werden sowohl männliche wie weibliche Gameten gebildet, oder sie können getrenntgeschlechtlich sein (diözisch), d. h., ein Thallus bildet nur einen Gametentyp aus. Bei Diözie erfolgt die Geschlechtsbestimmung genotypisch, d. h., sie wird genetisch durch eine entsprechende Chromosomenverteilung bestimmt. Bei Monözie wird die Anlage der Geschlechtsorgane durch andere, äußere Faktoren gesteuert, sie ist modifikatorisch (phänotypisch). – Bezüglich ihrer Individualentwicklungszyklen (Ontogenie) sind die Algen Haplonten oder Diplo-Haplonten, d. h. Algen mit einem Generationswechsel, seltener Diplonten. Diese Ontogenien sind bestimmt durch die zeitliche und räumliche Aufeinanderfolge der beiden Teilprozesse der sexuellen Fortpflanzung, Syngamie und Meiose. Die ursprünglichere Ontogenieform ist sicher die der Haplonten. Hierbei durchläuft der Organismus seine vegetative Phase in der Haplophase (n), also mit einfachem Chromosomensatz. Die Gameten entstehen mitotisch, und nach erfolgter Syngamie bildet sich die Zygote als einzige diploide (2n) Entwicklungsphase innerhalb der Ontogenie aus. Die Zygote entwickelt sich nach Ablauf der Meiose weiter, in der Regel unter Bildung von (n) Sporen. Haplonten sind viele einzellige Algen und viele Grünalgen (sowie einige Niedere Pilze). Diplonten sind im Pflanzenreich selten (z. B. die Diatomeen oder bei den Pilzen Saprolegnia). Bei den Diplonten wächst die Zygote mitotisch zu einem (2n) Organismus aus, der in Verbindung mit der Meiose Gameten bildet, die zur Zygote fusionieren. Hierbei sind in der Gesamtontogenie lediglich die Gameten haploid (n). Die meisten Algen (wie Pilze und alle Höheren Pflanzen) sind Haplo-Diplonten, d. h., während ihrer Individualentwicklung werden zwei (oder drei, Rotalgen) Generationen ausgebildet, die unterschiedliche Fortpflanzungskörper ausbilden (Generationswechsel). Allgemein wechselt sich eine haploide (n) Gametophyten- mit einer diploiden (2n) Sporophyten-Generation ab. Die beiden Teilschritte der sexuellen Fortpflanzung sind hier zeitlich und räumlich getrennt. Nach Syngamie der von Gametophyten abgegebenen Geschlechtszellen wächst die Zygote zum Sporophyten aus, auf dem in Sporangien, in Verbindung mit der Meiose, Sporen gebildet werden, die sich zum Gametophyten weiterentwickeln. Dieser "Generationswechsel" ist bei Pflanzen allgemein mit einem Kernphasenwechsel (n, 2n) verbunden (heterophasischer Generationswechsel). Die beiden Generationen können gleichgestaltet (isomorpher Generationswechsel) oder unterschiedlich gebaut sein (heteromorpher Generationswechsel). Häufig sind die morphologischen Unterschiede so groß, daß Sporophyten und Gametophyten als verschiedene Arten beschrieben wurden. Die Gametophyten sind gemischtgeschlechtlich (monözisch) oder getrenntgeschlechtlich (diözisch). Das häufig hierfür verwendete Begriffspaar "isothallisch–heterothallisch" ist irreführend.
Verbreitung und Vorkommen: Knapp 71% der Erdoberfläche sind von Meeren bedeckt; von dem verbleibenden Festlandteil entfallen ca. 3% auf freie Wasserflächen, Schnee- oder Eisfelder. Ein Vergleich der Verteilung der Arten zeigt, daß im Meerwasser vor allem die Braunalgen und Rotalgen sowie die Dinophyceae (Pyrrhophyceae, Feueralgen) und Chrysophyceae vorherrschen, während im Süßwasser und auf dem Festland die Grünalgen und Xanthophyceae verbreitet sind ( vgl. Tab. ). Algen können nahezu überall auf der Erdoberfläche frei schwimmend oder festsitzend vorkommen, sowohl in perennierenden wie auch auf nur zeitweise feuchten Standorten, in Trockengebieten, in heißen Quellen wie auf Eis und Schnee. Nach ihren Lebensansprüchen und ökologischen Standortbedingungen kann man sie in folgende Formenkreise gliedern:
a) Plankton: ein Sammelbegriff für alle im Wasser passiv treibenden Organismen; dazu gehört auch das Phytoplankton, aus einzelligen oder koloniebildenden Algen: im Meer besonders Kieselalgen und Vertreter der Peridiniales. Hauptsächlich Kalkflagellaten und Arten der Chrysophyceae bilden das Nanoplankton. b) Neuston: darunter versteht man meist einzellige Algen, die unter Ausnutzung der Oberflächenspannung des Wassers häufig nur zeitweise in oder auf der Wasseroberfläche leben, z. B. Euglena-Arten und Chromulina rosanoffi (Ochromonadaceae); leben sonst planktisch. c) Benthos (Benthal): hierzu gehören alle am Grund oder an untergetauchten Gegenständen festsitzende Algen, z. B. im Meer die ausgedehnten Bestände der Braunalgen und Rotalgen und im Süßwasser die von den Charales gebildeten Charawiesen. d) Algen der Fließgewässer, sind eine Lebensgemeinschaft benthischer und auch planktischer Algen, die in Fließgewässern unter anderem charakteristische Biozönosen bilden. e) Aerophyten (aerophytische Algen): außerhalb des Wassers an feuchten Standorten an Land lebende sog. Luftalgen, die Wasser aus der Atmosphäre aufnehmen und vor allem an der Schattenseite von Felsen und Bäumen bzw. im Boden (Bodenalgen; Bodenorganismen) leben. In den feuchten Tropen sind sie besonders häufig und siedeln dort auch auf Blättern. f) Thermale Algen (Thermalalgen), unter anderem in heißen Mineralquellen (Temperaturen zum Teil über 50 °C) lebende Algen. g) Kryophyten, Algen, die auf Schnee und Eis leben. h) Epibionten, Algen, die auf anderen Pflanzen (epiphytisch, z. B. Vertreter der Achnanthaceae) oder auf Tieren (epizooisch) festgewachsen leben. i) Symbiontische Algen, innerhalb anderer Organismen lebende Algen: hierzu zählen zum einen die in Flechten mit Pilzen in Symbiose lebenden Algengruppen, so z. B. Vertreter der Gattungen Chlorella (Oocystaceae) und Trentepohlia. Zum anderen gibt es endosymbiontisch lebende Algen in Form der Zooxanthellen auch in marinen Proto- und Metazoen.
Die wirtschaftliche Bedeutung der Algen ist erheblich größer als allgemein angenommen wird. Die Plankton-Algen sind die wichtigsten Primärproduzenten für energiereiche, organische Substanzen (Reservestoffe) in den Nahrungsketten der Meere und Binnenseen (Algenfresser). Algen haben eine wesentliche Bedeutung für die biologische Abwasserreinigung (Selbstreinigung) und dienen in diesem Zusammenhang auch der Charakterisierung der Gewässergüte (Saprobiensystem). Durch Eutrophierung ausgelöste Wasserblüten (Algenmatten, Algizide) können beträchtliche Schäden in der Fischereiwirtschaft und im Tourismus bewirken. Die Massenentwicklung von Algen kann mit der einhergehenden Bildung von Algengiften auch die Gesundheit des Menschen gefährden. Schon seit alters her werden unter anderem Meeresalgen wirtschaftlich genutzt. Einige Arten werden sogar für die menschliche Ernährung (Algenkulturen, Meereswirtschaft) und als Baumaterial (Algilit) genutzt. Braunalgen finden Verwendung als Futterzusatz und Dünger sowie bei der Iod- und Soda-Gewinnung. Von großer wirtschaftlicher Bedeutung sind die Phycokolloide aus den Zellwänden der Braunalgen (Alginsäure) und Rotalgen (Agar-Agar, Carrageenan). Weiter werden Kieselalgen, Grünalgen, Rotalgen und Braunalgen wirtschaftlich genutzt (Astaxanthin). Da einige Algen in extrem CO2-haltigem, saurem, alkalischem oder schadstoffbelastetem Milieu vorkommen können, wird sogar eine Nutzung bei der Rauchgasreinigung erprobt. – Wie viele Gefäßpflanzen sind auch zahlreiche Algen oligo- bis mesotropher Lebensräume durch die anthropogene Eutrophierung gefährdet und werden in Roten Listen (Rote Liste) geführt. Bei den gut untersuchten Zieralgen (Desmidiaceae) z. B. sind von den ca. 500 in Deutschland bekannten Arten fast zwei Drittel bestandsbedroht. – Agardh (K.A.), Bary (H.A. de), Ehrenberg (C.G.), Ingenhousz (J.), Naegeli (C.W. von), Oltmanns (F.), Pringsheim (N.), Trembley (A.); Algen I
Algen II
Algen III
Algen IV
Algen V
R.B./A.Se.
Lit.: Akatsuka, J.: Biology of economic algae. The Hague 1994. Bhattacharya, D. (ed.): Origins of Algae and their Plastids. Plant Systematics and Evolution, Suppl. 11. New York – Wien 1997. Ettl, H., Gärtner, G.: Syllabus der Boden-, Luft- und Flechtenalgen. Stuttgart 1995. Ettl, H.: Grundriß der allgemeinen Algologie. Stuttgart 1980. Margulis, L., Corliss, J.O., Melkonian, M., Chapman, D.J. (ed.): Handbook of Protoctista: the Structure, Cultivation, Habitats and Life Histories of the Eukaryotic Microorganisms and their Descendants Exclusive of Animals, Plants and Fungi. Boston 1990. Reitz, M.: Die Alge im System der Pflanzen. Stuttgart 1986. van den Hoek, C., Jahns, H.M., Mann, D.G.: Algen. Stuttgart 31993.
Algen
Farb- und Reservestoffe der Algen
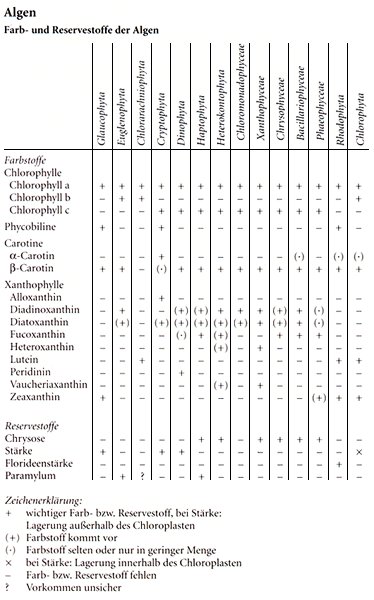
Zeichenerklärung:
+ wichtiger Farb- bzw. Reservestoff, bei Stärke:
Lagerung außerhalb des Chloroplasten
(+) Farbstoff kommt vor
(·) Farbstoff selten oder nur in geringer Menge
× bei Stärke: Lagerung innerhalb des Chloroplasten
– Farb- bzw. Reservestoff fehlen
? Vorkommen unsicher
| |
;) |
Algen
Vorkommen und relative Verteilung der wichtigsten Algen-Abteilungen bzw. -Klassen
| ||||
| Euglenophyta (Schönaugengeißler) | 800 | + + + + | + | |
| Chlorarachniophyta | 100 | – | + + + + | |
| Cryptophyta (Schlundgeißler) | 100 | + | + | |
| Dinophyta (Panzergeißler) | 2000 | + | + + + + | |
| Heterokontophyta Bacillariophyceae | 20 000 | + + + | + + + | |
| (Kieselalgen) | ||||
| Phaeophyceae | 1500 | (+) | + + + + | |
| (Braunalgen) | ||||
| Chrysophyceae | 1000 | + + | + + + + | |
| (Goldalgen) | ||||
| Xanthophyceae | 600 | + + + + | + | |
| (Gelbgrünalgen) | ||||
| Rhodophyta | 5000 | + | + + + + | |
| (Rotalgen) | ||||
| Chlorophyta | 8250 | + + + + | + | |
| (Grünalgen) |







Schreiben Sie uns!